The rapid expansion of the global centenarian population has shifted the focus of biogerontology from merely extending lifespan to decoding the molecular architecture of extended healthspan. Centenarians, semi-supercentenarians (105–109 years), and supercentenarians (≥110 years) possess a biologically distinct immune profile that delays or entirely avoids major age-related pathologies, such as cardiovascular disease and aggressive malignancies.
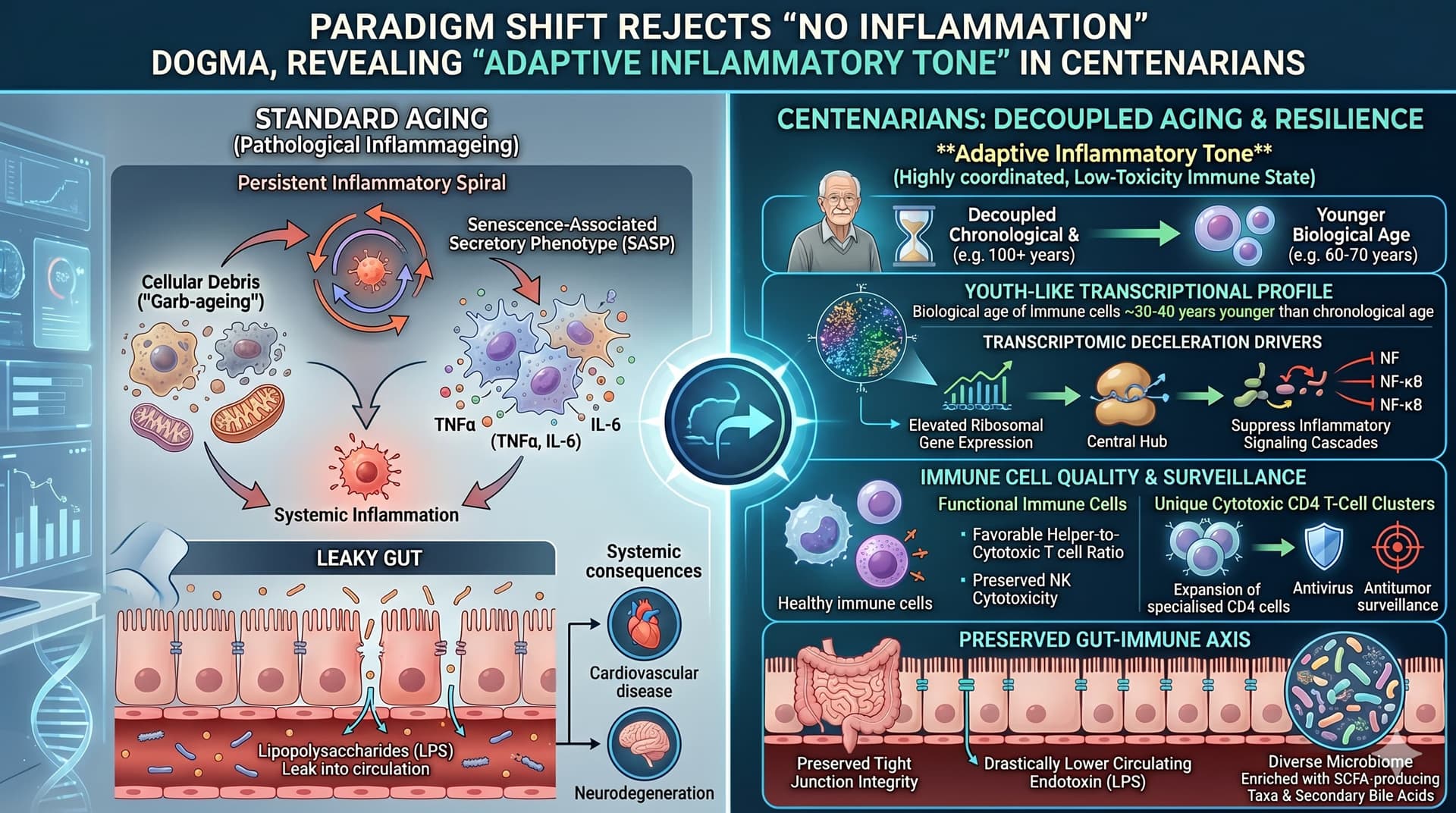
The primary paradigm shift established by this research is the rejection of the idea that exceptional longevity requires an absolute absence of systemic inflammation. Instead, long-lived individuals exhibit a highly coordinated, low-toxicity immune state termed adaptive inflammatory tone. While standard aging populations experience a destructive, feedforward inflammatory spiral driven by cellular debris (“garb-ageing”) and the senescence-associated secretory phenotype (SASP), centenarians effectively decouple chronological age from pathological inflammageing.
At the single-cell level, this adaptation is characterized by a youth-like transcriptional profile where the biological age of circulating immune cells can track up to 30–40 years younger than the individual’s chronological age. This transcriptomic deceleration is heavily driven by elevated ribosomal gene expression, which acts as a central hub to suppress inflammatory signaling cascades. Structurally, centenarians maintain a favorable helper-to-cytotoxic T cell ratio, preserve the functional cytotoxicity of natural killer (NK) cells, and display a unique expansion of highly specialized cytotoxic CD4 T cell clusters to maintain robust antiviral and antitumor surveillance.
Furthermore, this systemic resilience is structurally anchored by a preserved gut-immune axis. While typical aging degrades the intestinal lining—causing a “leaky gut” that leaks lipopolysaccharides (LPS) into circulation to fuel systemic inflammation—centenarians maintain tight junction integrity, drastically lower circulating endotoxin levels, and maintain a highly stable, diverse microbiome enriched with short-chain fatty acid (SCFA)-producing taxa and protective secondary bile acids. Ultimately, centenarian longevity is not a passive absence of defects, but an active, multi-layered physiological calibration that maintains homeostatic equilibrium under lifelong antigenic stress.
Actionable Insights for Longevity
-
Suppress the NLRP3 Inflammasome: Healthy centenarians maintain low, youth-like expression of the NLRP3 inflammasome in T cells, a state that correlates with delayed bone loss, reduced frailty, and extended lifespan. Targeted interventions should focus on pharmacological or dietary strategies (such as periodic fasting or ketone body production) to suppress baseline NLRP3 activation.
-
Upregulate Autophagy Pathways: Centenarians display elevated circulating levels of Beclin-1 and downregulate tissue hormones like acyl-coenzyme A-binding protein (ACBP) that inhibit autophagy. Maintaining robust autophagy-lysosomal flux via caloric restriction mimetics (e.g. rapamycin) or targeted compounds suppresses SASP and shields immune cells from age-associated dysfunction.
-
Enrich SCFA-Producing Gut Microflora: Exceptional longevity tracks with high abundance of Akkermansia muciniphila , Bifidobacterium , and the Christensenellaceae family. Prioritizing a diverse, plant-forward diet high in prebiotic fibers optimizes short-chain fatty acid (SCFA) production, preserves intestinal barrier integrity, and drops serum zonulin and LPS to eliminate systemic endotoxemia.
-
Mitigate Cytoplasmic Nucleic Acid Accumulation: Centenarians overexpress the RNAse H2 subunit RNASEH2C , which clears cytoplasmic RNA:DNA hybrids. Suppressing the build-up of these immunogenic fragments restrains the chronic activation of DNA damage-induced inflammatory pathways and type I interferon signaling.
Source:
- Paywalled Paper: The long-lived immune system of centenarians
- Journal Name: Nature Reviews Immunology.
- Key Institutional Affiliations: University of California Irvine (USA), Universidad Europea de Madrid (Spain), University of Oviedo (Spain).
- Geographic Origin: International Collaboration (primarily United States and Spain).
- Impact Evaluation: The impact score of this journal is 108.3, evaluated against a typical high-end range of 0–60+ for top general science, therefore this is an Elite impact journal.