Speaking of NMR consuming Tubers(i.e. root veggies such as sweet potatoes) and subterranean low sunlight vitamin D independenr metabolic pathways:
Eating tubers helps naked mole rats achieve exceptional longevity primarily through their effects on metabolism, nutrient intake, and cellular health���. Tubers are low-protein, low-methionine plant foods, and naked mole rats have adapted to thrive on this diet, which closely resembles the dietary interventions that extend lifespan in other model organisms��.Nutritional Composition and Metabolic EffectsTubers are rich in complex carbohydrates and fiber, but low in protein and particularly low in the amino acid methionine��.
Naked mole rats show plasma profiles with low levels of circulating methionine, a feature that mimics the benefits seen in methionine-restricted diets proven to extend lifespan in mice, rats, and other animals�.Their diet supports a low metabolic rate and helps prevent age-related metabolic decline, contributing to the maintenance of stable physiological functions well into old age��.
Impact on Aging and Health: Methionine restriction from tuber diets is linked to lower oxidative stress, resistance to cancer, and delayed onset of age-related diseases in naked mole rats��.Naked mole rats retain youthful metabolic profiles throughout their lives, similar to animals on caloric restriction or extended fasting periods�
Their natural underground diet of tubers also promotes a stable gut microbiome, further supporting health and longevity�.
Several compounds in tubers are linked to enhanced DNA repair and longevity, both via direct biochemical action and through antioxidant effects that protect DNA from damage.
Key Tuber-Derived Compounds
Caffeoylquinic acids (chlorogenic acids): Potent antioxidants found in many tubers that stimulate cellular repair of oxidative DNA damage, helping maintain genome integrity.
Phenolics and flavonoids: Broadly present in tubers such as potatoes, sweet potatoes, and yams, these compounds scavenge free radicals and support excision repair mechanisms, decreasing mutation rates and protecting cells against aging.
Beta-carotene and Vitamin C/E: Vitamins abundant in tubers work as antioxidants, reducing oxidative stress and aiding enzymatic DNA base repair, thus contributing to longevity.
Terpenoids and glycosides: Select tubers harbor bioactive terpenoids and glycosides found to extend lifespan in animal models, partially by upregulating genes involved in stress resistance and DNA repair.
Mechanisms Promoting LongevityAntioxidant compounds reduce DNA damage from metabolic processes and environmental stress.
Some tuber-derived molecules actively stimulate cellular DNA repair pathways, including base excision and homologous recombination, crucial for maintaining stable genetic function throughout aging.
These compounds result in a lower incidence of cancer and age-related diseases, as seen in animal models and suggested by naked mole rats’ exceptional lifespan.
An Okinawa diet!!
Nick this is very interesting. Can you share what the sources are for these two posts above?
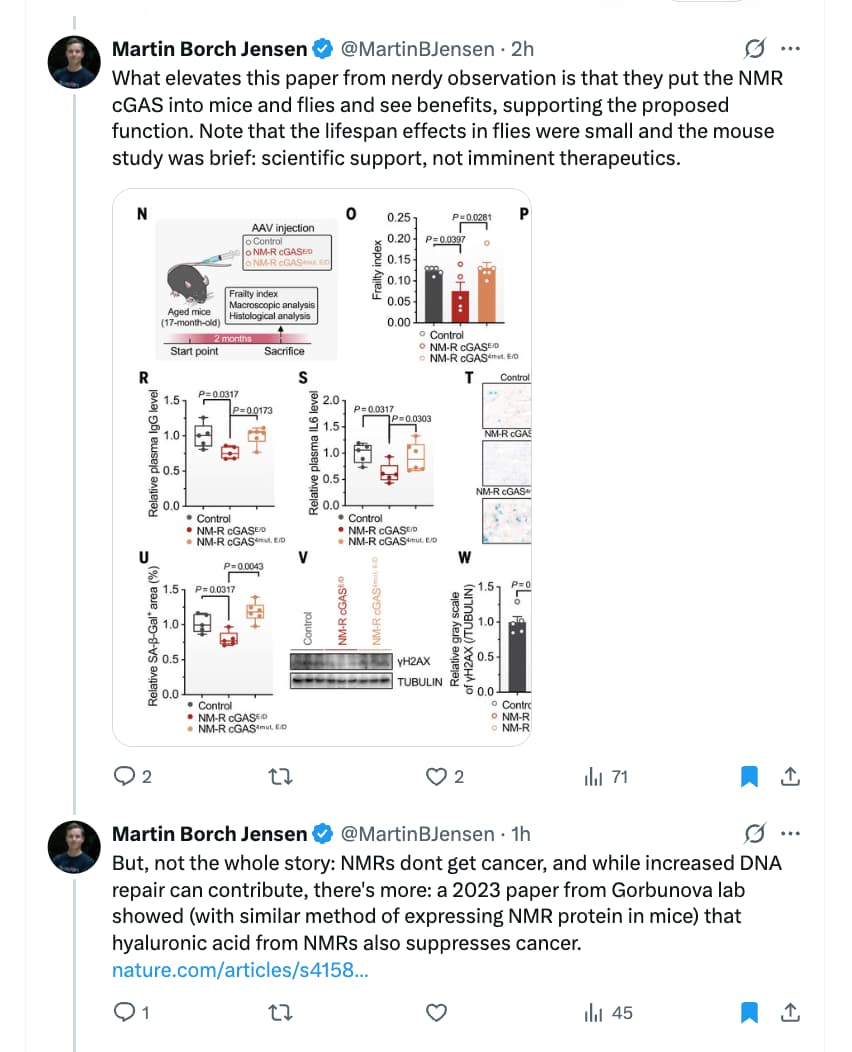
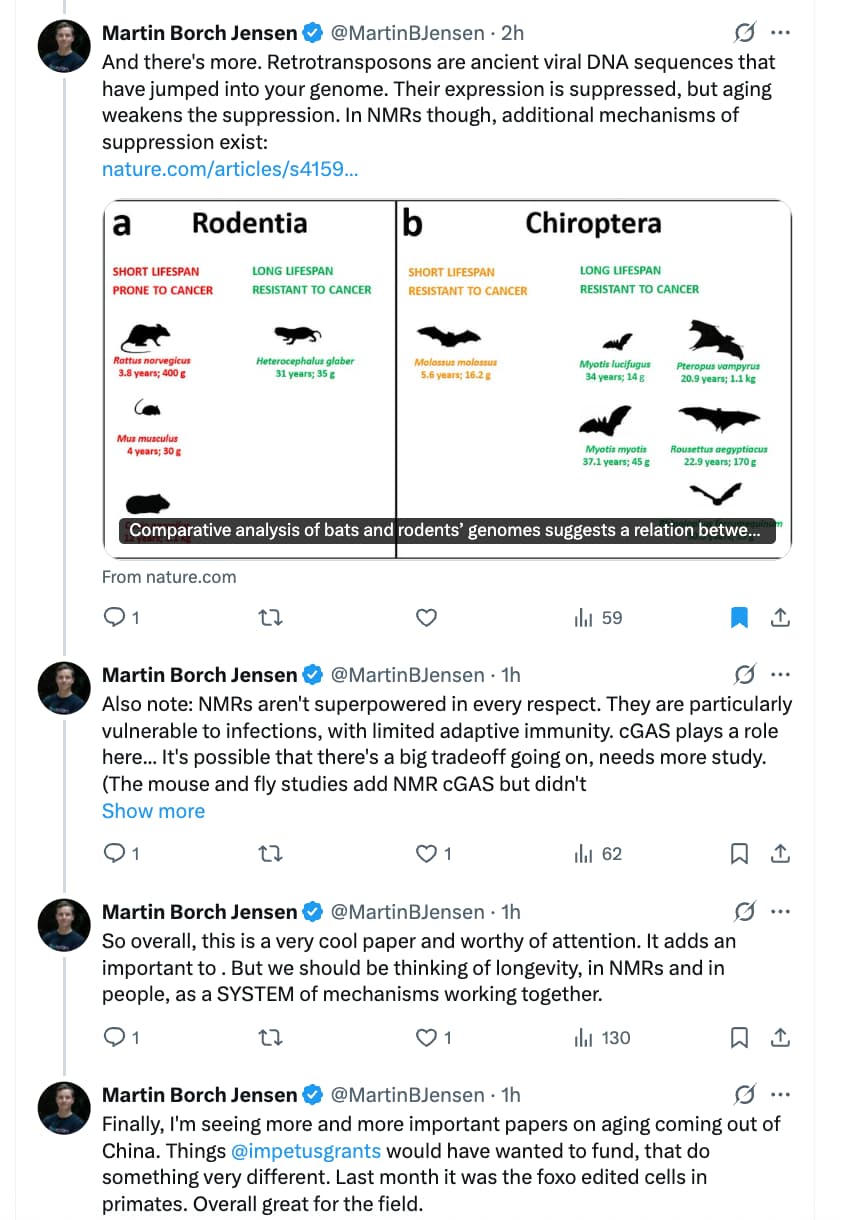
Source: https://x.com/MartinBJensen/status/1976660387111342160
Papers:
If you’re interested in the science of healthspan and longevity, you’ve likely heard of the naked mole rat, a remarkable outlier among mammals that defies the laws of biologic aging. For nearly 30 years, we didn’t know what accounted for this phenomenon, but a compelling new report sheds light on a pathway that was only discovered in 2012, awarded the Lasker Prize in 2024, and is now the target of many clinical trials. This edition of Ground Truths is about the convergence of two exciting life science discoveries and how they may influence the future of medicine.
Background: Two Threads of Life Science Converge
By the late 1990s it was noted that there was something unusual about the lifespan of naked mole rats. In 2002, Drs. Rochelle Buffenstein and Jennifer Jarvis, comparative biologists, published “ A New Record for the Oldest Living Rodent” for the naked mole rat, living over 27 years, about 5 times the maximal lifespan of mice or rats. In the past 2 decades since this discovery, much work done to understand their longevity and remarkable resistance to age-related diseases. Their heart, kidneys, liver, eyes and bones remain youthful. Cognitive function is preserved and they do not develop cancer (only 5 cases in over 3,000 necropsies). Some have lived up to 39 years of age. It has been shown that the naked mole rat lifespan defies Gompertz’s law for not having an exponentially increasing risk of death as they get older—the naked role rat is considered a “non-aging mammal.” Their existence is almost completely underground, moving around in tunnel systems. They are certainly not attractive! (Figure below) They are hairless, buck-toothed, wrinkly, and can survive without oxygen for 18 minutes. Known as extreme xenophobes, they only seem to die when they beat each other up. Owing to the unique features of healthspan and lifespan, they are considered the supermodel organism for understanding these critical biological features.
But until now, we didn’t know the underlying mechanism for how naked mole rats lived so long and so healthily. This week, in Science, Yu Chen and colleagues from China published a compelling case for what explains the naked mole rat story.
Before we get to that, let’s go to a major basic science discovery that was awarded the 2024 Lasker Prize (considered pre-Nobel). Last year’s basic medical research prize recipient was Dr. Zhijian Chen of the University of Texas, Southwestern, for the discovery of cGAS (cyclic guanosine monophosphate-adenosine monophosphate synthase (yes a mouthful, thank goodness for acronyms). In back to back papers in Science in 2012 and 2013, Chen and colleagues identified and described cGAS and the pathway. It serves as a danger sensor, predominantly present in the cell cytoplasm, detecting penetration of foreign DNA from a virus or bacteria, or damaged DNA in the cell. Once danger is sensed, cGAS is activated and through cGAMP there is activation of STING, which stands for the stimulator of the interferon gene. This critical step leads to production of Type I interferons, a principal component of our innate immunity (simplified Figure below). Recall that with SARS-CoV-2 infection, Type 1 interferon, our first line of defense, provided critical protection, such that people with autoantibodies to type 1 interferon were highly vulnerable to severe, life-threatening Covid.
You can also see this pathway’s role in promoting inflammation, cell senescence, and death. Since the discovery in 2012, the cGAS-STING pathway has been implicated in many autoimmune and age-related inflammatory diseases, including cancer and neurodegenerative.
“Solved” seems optimistic. The claimed effects in mice seem impressive (without reading the paper, which I can’t access). On the other hand, there’s no lifespan claim for mice, possibly due to resource constraints.
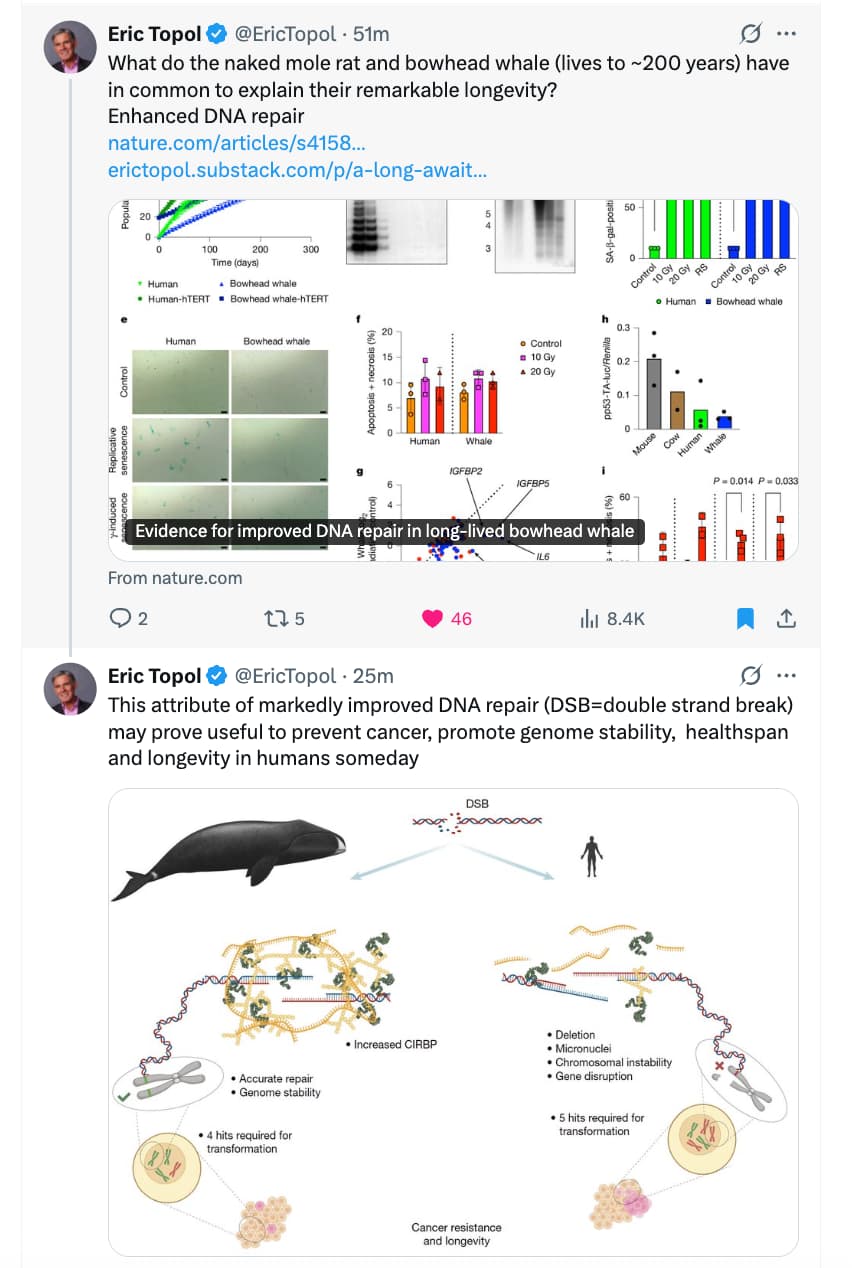
Source: https://x.com/EricTopol/status/1983577674846294078
Papers:
Evidence for improved DNA repair in long-lived bowhead whale
Is there anything we can do now to enhance DNA repair?
A little… but you’re probably already doing most of these things. I know of at least one company that is working on bringing drugs to market that protect DNA from double-strand breaks.
From CGPT5
Here is a prioritized summary of the major interventions we discussed for enhancing DNA repair/genome stability — ranked by strength of evidence, effect size (where reported), and relevance to healthy human populations. I’ll include for each: effect size (if known), evidence quality, key caveats, and a “priority score” (relative) to help you decide how to allocate effort/attention in your optimisation framework.
| Rank | Intervention | Reported Effect Size* | Evidence Quality & Quantity | Key Caveats | Priority Score† |
|---|---|---|---|---|---|
| 1 | Micronutrient optimisation (e.g., folate/B12/zinc/vitamins C/E/selenium) | Moderate but variable: e.g., one study showed increased vitamin C supply reduced 8‑oxodG ~2‑fold in sperm DNA under low‑intake conditions. | Fair: Several human observational & intervention studies, but many with mixed results, short durations, surrogate endpoints only. | Most data relate to deficiency→adequacy transitions, not supra‑normal supplementation. Effects differ by baseline status & exposure load. | ★★★★☆ |
| 2 | Polyphenol‑rich diet / phytochemicals(berries, anthocyanins, curcumin, other flavonoids) | Small to moderate: Review shows consistent reductions in DNA damage markers in human diet studies (Comet assay) but effect sizes often modest and heterogeneous. | Moderate: Many human dietary interventions (though few RCTs), many cell/animal studies. | Bioavailability issues, dose/compound heterogeneity, many studies in healthy young adults with low baseline damage load. | ★★★☆☆ |
| 3 | Sleep / circadian alignment (ensuring proper sleep to support DNA repair) | Indirect effect, but mechanistically strong: studies show sleep deprivation increases DNA damage and reduces repair gene expression. | Good mechanistic evidence, less human RCTs for “sleep optimisation → DNA repair” endpoint. | Hard to quantify “how much better” repair gets; likely foundational rather than “boost” only. | ★★★★☆ |
| 4 | Melatonin supplementation | In one human RCT (40 night‑shift workers), 3 mg melatonin for 4 weeks led to ~1.8‑fold increase (95% CI 1.0‑3.2) in urinary 8‑OH‑dG (marker of oxidative DNA damage repair) during day sleep. p ≈0.06 (borderline). | Moderate but limited: Only one small RCT in a specific population (night‑shift workers). Many other mechanistic/cell studies. | Specific population (shift workers) with suppressed melatonin; effect in general healthy population unclear. Dose/long‑term safety/efficacy unknown. | ★★☆☆☆ |
| 5 | Targeted DNA‑repair drugs / advanced agents (e.g., PARP activators, NAD+ precursors, sirtuin modulators) | Preclinical only (cell & animal) showing enhancement of repair pathways (e.g., NAD+ dependent PARP activity) but no robust human trial data for healthy individuals. | Weak: mechanistic data strong, human outcome data basically absent. | Unclear safety, dose, long‑term effects; potential off‑target/unintended consequences. | ★☆☆☆☆ |
- “Effect size” refers to measured changes in DNA‑damage/repair surrogate markers (e.g., 8‑OH‑dG, comet assay) rather than hard endpoints (cancer incidence, longevity)
† Priority Score: ★☆☆☆☆ = lowest, ★★★★☆ = higher priority
Recommendations—How to use this ranking in your optimisation framework
- Top priority interventions: Micronutrient optimisation + sleep/circadian hygiene. These have the best evidence, are broadly applicable, relatively safe, and fit your existing biomarker/lifestyle infrastructure.
- Intermediate priority: Polyphenol‑rich diet/phytochemicals. Because effect sizes are smaller and more inconsistent, but the cost/risks are low and fit into a “supportive” layer.
- Lower priority: Melatonin supplementation (for general population) and advanced DNA‑repair drugs/agents. These may be useful in specific high “burden” scenarios (e.g., night shift work, high oxidative stress exposure, chronic inflammation) but are less generalisable to healthy optimisation without more human data.
Additional nuance & caveats you should integrate given your deep biomarker focus:
- Many studies measure DNA damage reduction (less insult) rather than enhancement of repair capacity per se. So effect may be from less new damage rather than faster repair.
- Baseline status matters a lot: individuals with micronutrient deficiencies, high DNA damage burden (older age, heavy exposures) show larger effects; healthy optimised younger individuals likely show smaller incremental gains.
- Surrogate markers dominate (8‑OH‑dG, comet assay tail moments). We do not yet have robust RCTs showing that modest improvements in those markers translate to lower cancer risk or longer lifespan.
- There may be diminishing returns (once you reach “adequate” micronutrient status, further supplementation may produce minimal repair/benefit but potential risk).
- Interactions matter: e.g., antioxidant supplementation might blunt adaptive stress responses; repair pathways may be inhibited if you reduce oxidative “signals” too much (a hormesis perspective) — although specific human data are lacking.
- Long‑term safety/optimal dosing remain uncertain for many phytochemicals/novel agents.
Life Lessons from (Very Old) Bowhead Whales
A gene that helped bowheads adapt to frigid Arctic waters also granted them extraordinary longevity. Could it help aging humans become more resilient?
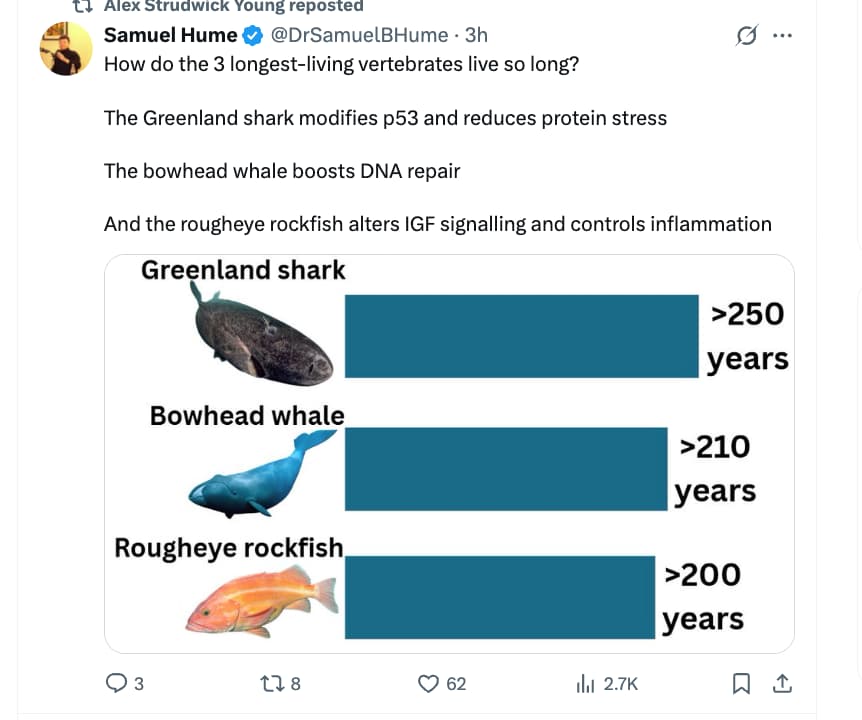
For more than 1,000 years, the Inupiat people of Alaska have hunted bowhead whales in the Arctic Ocean. Over the centuries, they grew to appreciate the long lives of the animals, the longest-living mammals on Earth. Generations of hunters could recognize the same individual at sea. Inupiat captains have told researchers that a bowhead whale lives two human lifetimes.
Scientists now suspect that bowheads can live even longer than that. Some whales caught in the late 1900s had old harpoon points lodged in their blubber that dated to the mid-1800s. By measuring the molecular damage that accumulates in the eyes, ears and eggs of bowhead whales, researchers have estimated that bowheads live as long as 268 years.
A study published in the journal Nature on Wednesday offers a clue to how the animals manage to live so long: They are extraordinarily good at fixing damaged DNA.
The new study was led by Vera Gorbunova and Andrei Seluanov, a married couple who both work at the University of Rochester and study long-lived mammals such as bats, beavers and naked mole rats, along with bowhead whales. They and their colleagues are uncovering many molecular adaptations that extend animal lives. These species, the research shows, are gaining years thanks to increases in the levels of certain proteins and subtle changes in how these proteins work with others.
“We’re not talking about new genes,” Dr. Seluanov said. That finding raises the possibility that similar changes could be reproduced in humans to extend our own healthy life span. “We need to just tweak our system a little bit to resemble what we’ve found in naked mole rats or bowhead whales,” Dr. Seluanov said.
As it turns out, bowhead whale cells produce large amounts of a protein called CIRBP. Its job is to speed up the production of other proteins that protect against cold-triggered damage to cells.
Dr. Gorbunova and Dr. Seluanov also noticed a lot of CIRBP floating around the whale’s DNA as well. They found a single study, published in 2018, suggesting that CIRBP might also help repair DNA. Indeed, when Dr. Gorbunova and Dr. Seluanov inserted the bowhead CIRBP gene into human cells, the rate of DNA repair in those cells doubled.
The DNA-fixing protein, and the gene that produces it, appear to be key to the bowhead’s longevity. Over an animal’s lifetime, damaged DNA builds up throughout its body, leading to many ailments, not just cancer. When the scientists engineered the bowhead’s CIRBP gene into fruit flies, those flies lived longer than those with the normal insect version of the gene.
Read the Full story: Life Lessons from (Very Old) Bowhead Whales (NY Times)
And we can focus on preventing DNA damage…
Above ChatGPT claims that increased urinary 8‑oxodG is a marker of increased DNA repair but I think that’s misleading. Yes increased 8‑oxodG in the urine is a marker of more DNA having been repaired, but that doesn’t necessarily indicate improved DNA repair. It might as well just mean there was more DNA to repair rather than your body having been repairing more of the damage that is present. As an example, if your DNA damage remains constant but your body suddenly starts repairing 50% more of it, then urinary 8‑oxodG would increase about 50%. However if you suddenly have 50% more DNA damage but are repairing the same proportion of the damage as before, then urinary 8‑oxodG would also increase by about 50%. But in the latter case, the increase isn’t reflecting improved repair but more damage.
This YAK study looking at CIRBP is worth a look:
https://www.mdpi.com/2218-273X/15/6/759
shows that rapamycin and CIRBP share common mechanisms
Essentially: Yaks “over-express” CIRBP for cold adaptation. The study used Rapamycin as a “positive control” Finding: if they knock down the Yak’s CIRBP expression then Rapamycin will come to the rescue and replicate the lost CIRBP benefits
Per Grok 4.1
Summary of the Study
Title: CIRBP Enhances the Function of Yak Cumulus Cells by Activating AMPK/mTOR-Mediated Mitophagy
Authors: Rui Zhang, Yan Cui, Yangyang Pan, et al.
Journal: Biomolecules (Volume 15, Issue 6, Article 759)
Publication Date: 24 May 2025
DOI: 10.3390/biom15060759 (Open access; full text available on MDPI and PMC)
Background and Rationale
- Yaks (Bos grunniens) live in high-altitude, cold, hypoxic environments on the Qinghai-Tibet Plateau.
- Cumulus cells (surrounding the oocyte in the follicle) are critical for oocyte maturation, steroid hormone production, cumulus expansion, and protection against apoptosis.
- Cold-inducible RNA-binding protein (CIRBP) is a stress-response protein upregulated by mild hypothermia (e.g., 32 °C), hypoxia, and other stressors.
- The researchers hypothesized that CIRBP helps yak cumulus cells adapt to cold/hypoxic stress via regulation of mitophagy (selective autophagy of damaged mitochondria).
Key Methods
- Yak cumulus-oocyte complexes (COCs) were cultured at normal (38.5 °C) vs. mild hypothermic (32 °C) temperatures.
- CIRBP was overexpressed (via lentiviral vector) or knocked down (siRNA) in primary yak cumulus cells (YCCs).
- Mitophagy was manipulated using:
- Inhibitors: Mdivi-1 (mitochondrial fission/mitophagy inhibitor) and 3-methyladenine (3-MA, general autophagy inhibitor).
- Activators: Rapamycin (RAPA; direct mTOR inhibitor, used as a positive control) and AICAR (AMPK activator).
- Outcomes measured: steroid hormone secretion (estradiol E2, progesterone P4), cumulus expansion, apoptosis rates, mitochondrial function, mitophagy markers (e.g., LC3-II, PINK1/Parkin pathway), and signaling proteins (p-AMPK, p-mTOR, etc.).
Main Findings
-
Mild hypothermia (32 °C) mimics cold stress:
- Increased CIRBP expression.
- Activated autophagy/mitophagy.
- Enhanced E2/P4 secretion and cumulus expansion.
-
CIRBP overexpression:
- Upregulated mitophagy-related proteins (LC3-II, PINK1, Parkin, etc.).
- Activated AMPK (↑ p-AMPK) → inhibited mTOR (↓ p-mTOR) → induced mitophagy.
- Increased E2 and P4 secretion, promoted cumulus expansion, reduced apoptosis.
- Rescued cells from dysfunction caused by mitophagy inhibitors (Mdivi-1 or 3-MA).
-
Rapamycin (mTOR inhibition) as positive control:
- Produced nearly identical beneficial effects to CIRBP overexpression (enhanced mitophagy, hormone secretion, anti-apoptosis).
- Confirms that the protective effects converge on mTOR suppression and mitophagy activation.
-
Pathway confirmation:
- CIRBP primarily acts via AMPK activation → mTOR inhibition (not PI3K/AKT).
- Blocking mitophagy abolished CIRBP’s benefits.
Conclusions
- CIRBP is an endogenous stress-adaptive protein in yak cumulus cells that improves cellular function (hormone production, expansion, survival) under cold/hypoxic conditions by inducing protective mitophagy via the AMPK/mTOR pathway.
- This represents a natural mechanism that partially mimics the effects of rapamycin (a pharmacological mTOR inhibitor and known autophagy inducer).
- Potential relevance: Insights into cold adaptation in plateau animals; possible implications for therapeutic hypothermia, reproductive biology in stressful environments, or longevity pathways (mitophagy/mTOR are conserved anti-aging targets).
This is currently the only published study directly linking CIRBP to mitophagy regulation in any cell type, and it highlights a novel “rapamycin-like” endogenous pathway.
Some animals have a very different relationship to aging than we do: They don’t get cancer, they never go through menopause, and they live absurdly long lives.
For instance, one bat species can live for more than 40 years, which may not sound like very long but that’s about nine times longer than expected based on its size. For comparison, if we aged on that scale, we’d live for hundreds of years. These bats aren’t the only animal super-agers—there’s a whole menagerie of them.
So what’s their secret? And can we learn anything from them that might help us live longer, healthier lives? Host Flora Lichtman talks with longevity researchers Vera Gorbunova and Juan Manuel Vazquez about what animals are teaching us.
Guests:
Dr. Vera Gorbunova is a biologist and professor at the University of Rochester, and a co-director of the Rochester Aging Research Center.
Dr. Juan Manuel Vazquez is a biologist and assistant professor at Pennsylvania State University studying the evolution of aging.
Transcript: Podcast transcripts, sponsors, and audience data - Podscribe
Excerpt from Transcript:
4
00:08:30
Well I must say a word about elephants so that you will understand.
0
00:08:34
Okay, okay.
4
00:08:35
Okay. Because in the elephants, they enhanced the way cells are the surveillance. So like eliminating cells that are damaged. So, so that is the strategy for elephants. Bad cells get rid of them very quickly. And that was our hypothesis for the whale. But this is not what we found. We found that the whale enhances maintenance, they just don’t accumulate mutations as fast. And when they are faced with DNA damage, they deal with it much more efficiently. They don’t kill their cells very readily, but they maintain their cells like they don’t let things deteriorate to the point that it’s necessary to eliminate the cell.
0
00:09:20
How do they do it?
4
00:09:22
Well they have very high levels of a protein called C-I-R-B-P or cold induced RNA binding protein. And what we found is that this protein promotes more efficient DNA repair and it protects from mutations. So we humans also have this protein, but we make very small amount of it and whales make maybe a hundred times more. Hmm. And the, as the name suggests, it’s cold deduced, so it has something to do with cold and whales live in coal. But we hypothesized that evolutionarily they upregulate this protein first to deal with cold, but then it was also coped to help the near repair.
4
00:10:08
And this way whales enhance their maintenance strategy. Because I’m thinking the strategy of an elephant is good, but maybe up to a point, if you plan to live more than a hundred years and you’re eliminating cells very readily, you may deplete your stem cell resource
0
00:10:26
Run outta cells. Yes.
4
00:10:29
So it may be for the lifespans that exceed 200 years, well you need a different strategy. You just need to prevent these mutations in the first place.
0
00:10:41
Did you, did you test putting this protein or, or sort of higher levels of this protein in human cells?
4
00:10:47
What happened? Yes. That was the most exciting part. When we put this protein in human cells, it improved the way cells repaired breaks in the DNA. So they became about twice more efficient. So just with one protein, which for me was the most exciting finding because it means there is room for improvement in human dary repair. Hmm. For many researchers, human dary repair seemed like something like okay, we are given it. We cannot make it any better if we even try, we can change the balance between different repair proteins and things would get worse.
4
00:11:28
But what this study demonstrates that we actually can make it better and that gives hope for longevity and also for cancer prevention. Because if we can make ourselves not generate mutations, we can prevent cancer from happening.
0
00:11:45
Manny, the bats you study also seem to not get cancer, right? Is the mechanism the same? If you
3
00:11:52
Look at the leaderboard for the 20 species of the lowest cancer risk, about five of them are bats. So bats both have the ability to improve their like DNA repair. So when they acquire damage, they’re better at dealing with it than like say a mouses. But they also have a very elephant like mechanism where they’re also able to purge damaged cells. So it’s basically having like both the ability to maintain a better genome and also a better sorting facility for getting rid of the jump.
Actionable Insights:
You know Vera, I think everyone listening to this is wondering can we learn from these animals to help ourselves live not just longer lives but healthier lives?
4
00:14:14
Oh yes. We could learn so much and if we talk about genome maintenance and more accurate D repair that seems to be conserved across many long lived mammals, long lived animals. But the mechanisms are different, which is amazing. So we looked at anti-cancer mechanisms in naked moat we find something entirely different. They evolved unique adaptations linked to subterranean lifestyle. They started making a lot of hyaluronic acid to squeeze through tunnels and that now prevents cancer in them.
0
00:14:51
Oh wow. And also people know that from skincare, right?
4
00:14:54
Yes it is. It’s skincare, it in many cosmetics, but in the naked bone they have it inside the skin in the very large quantities. So their evolution took this path and we humans, we also pretty good at DNA repair, but we didn’t have the same evolutionary pressure. So we, we are missing some of these adaptations and we can develop some strategies to bring these adaptations into human biology and then we would definitely benefit because it’s something that we are missing but we can improve upon.
0
00:15:28
How do you, what do you expect the treatment to look like? Is it gene therapy? Is it a pill of whe protein?
4
00:15:35
Well, there may be different strategies depending on every adaptation. So let’s say with naked Mole Rat, we are a bit, a little bit further along because we were thinking, okay, how can we increase the level of hyaluronic acid? Of course you can apply it with a cream, but it won’t get through your skin. It’s just a good moisturizer. But if we develop small molecules to slow down breakdown of the degradation of hyaluronic inside the skin, then we can increase our own levels of hyaluronic acid. And that was, we published a paper because we gave this small molecule to mice with cancer and they became a little bit more like naked rads that tumors didn’t spread it just with applying the small molecule that could be orally administered.
3
00:16:27
We also, because of basic science research, just have a lot of data and also pharmacological research, a lot of these big databases of how different drugs change your gene regulatory networks. So another way of translating this research is just looking at those two things with machine learning and taking advantage of the new AI tools now out to basically look for drugs that are already FDA approved that happen to have these like beneficial like, oh it makes this look more like the naked Mole rat or it makes this look more like the Bowhead whale. Right?
0
00:16:58
Is any of this moving out of the lab and into, you know, clinical trials?
4
00:17:03
Yes. Have one. We analyzed the innate pair across 25 species of rodents and we found in the innate pair was more efficient in long-lived rodents due to another protein, not CRBP, but protein called SIRT6. It’s also a genome maintenance protein and we search for activators and we found a natural compound from brown seaweed called Fucoidan. So it’s very healthy people in Japan. South Korea eat brown seaweed is part of soups and stews. So it’s, it’s very safe.
4
00:17:43
And we took Fucoidan and we found it activates SIRT6 very strongly. Then we gave Fucoidan to old mice, and mice started to live longer and their genome stability improved. So right now we already started a clinical trial and we see if now the strategy that we learned from rodent studies can actually be safely applied to extend human health.
0
00:18:08
Fucoidan. You heard it here first. Manny. What’s your hope for this research?
3
00:18:13
You know, if you ask an audience of people who here has had someone die of cancer, who here has had know someone who has like died of heart disease, right? So like you don’t usually get hands that are lowered and these are things that have a huge like societal impact, right? Like we spend so much money just trying to like play this game of whack-a-mole to deal with like all different ways cancer can pop up and all the different ways heart disease can pop up. The thing that’s like really interesting about studying long-lived animals is that they live long. And so as a result they have a lot of preventative mechanisms as well. So we’re not just talking about like using like nick and Mo rats and elephants and whales and bats to discover cures for cancer and aging and like heart disease.
3
00:18:58
We’re actually also talking about looking for preventatives for all these things. So that way people will eventually like be in a generation that barely knows what cancer is, right? Or you know, like any other kind of like agent related illness.
0
00:19:13
Vera, your hopes for this research.
4
00:19:16
Yes. I am also looking into the future where there will be less human suffering because these diseases, cancer, heart disease, diabetes, they cause so much human suffering. And if we learn from evolution how to live like a bot that can fly until its last day, I mean that, that would be amazing if people could enjoy their lives productively until the very last day. So this is my hope.
Related discussion thread here: Fucoidans (SIRT6 activators) from brown algae. Where to source?
A simplistic look at this research would suggest that if we just inserted extra copies of the P53 gene in humans, cancer would be a non-issue. Alas, not so simple:
ChatGPT5.1 Summary:
Short version:
Yes—people have made “extra-p53” mammals. In mice, extra Trp53 copies clearly lower cancer incidence, but by themselves they do not robustly extend lifespan. The only clear extension of lifespan comes from models with extra, but tightly regulated, Arf + p53 together, and even there the effect is modest. When p53 is made too active or deregulated, you get cancer resistance plus premature aging and shorter life.
1. Core mouse models with extra p53 copies
1.1 “Super p53” mice – extra Trp53 under its own promoter
Construct / design
- Bacterial artificial chromosome (BAC) transgene carrying an intact Trp53 locus (promoter + regulatory elements), added on top of the two endogenous alleles → many animals effectively have 3 functional copies of p53 (“super p53”). (PMC)
Cancer outcomes
-
Cells from super-p53 mice show:
- Higher basal p53 levels and stronger induction after DNA damage.
- More robust induction of p21 and increased apoptosis after irradiation. (PMC)
-
In vivo, they have markedly reduced spontaneous tumor incidence and delayed tumor onset versus wild-type littermates. (PMC)
-
Used later as a gain-of-function model to show that an extra p53 copy suppresses Kras-mutant lung adenomas and lymphomas, although it does not protect well against radiation-induced lymphomas. (JCI Insight)
Longevity / aging
- Original EMBO J paper: super-p53 mice age normally, with no obvious progeroid features. (PMC)
- No convincing increase in lifespan: survival curves are essentially superimposable on wild-type; ~70% of mice in both groups are alive at 2 years in one report. (Science)
- Follow-up work where super-p53 was combined with telomere dysfunction shows that increased p53 activity does not exacerbate telomere-driven degenerative aging, arguing that physiologically regulated extra p53 can be “safe” for aging. (EMBO Press)
Bottom line:
Extra, normally regulated p53 → stronger DNA-damage response and fewer cancers, but no lifespan extension and no premature aging.
1.2 Constitutively high p53 activity via Mdm2 attenuation
Construct / design
- Mendrysa et al. engineered mice with reduced expression of Mdm2, the main negative regulator of p53. This leads to chronically elevated p53 activity, somewhat analogous in outcome to “more p53,” even though the gene copy number itself isn’t increased. (PubMed)
Cancer outcomes
- These mice are highly cancer-resistant across several tumor models. (PubMed)
Longevity / aging
- Despite “constitutively high” p53 activity, they do not show accelerated aging and have essentially normal lifespan, contrasting with earlier hyperactive p53 mutants (see below). (PubMed)
Interpretation:
It supports the idea that moderate, well-controlled elevation of p53 activity can give cancer protection without obvious aging penalty—similar conclusion to super-p53—but again, no clear life-extension.
1.3 Extra Arf + p53 dosage – where you do see lifespan extension
These models are closest to what you asked about in terms of both cancer and longevity.
1.3.1 “Super Ink4a/Arf” alone
- Extra copy of the Ink4a/Arf locus, giving modestly higher p16^Ink4a and p19^Arf activity, under normal regulation. (arep.med.harvard.edu)
- Phenotype: increased cancer resistance, but normal aging and lifespan; no significant life extension. (Genes & Development)
1.3.2 “s-Arf/p53” (super Arf + super p53, both regulated)
- Matheu et al. then crossed “super p53” with “super Ink4a/Arf” to generate mice with extra, but normally regulated, copies of both p53 and Arf (often called s-Arf/p53). (Nature)
Cancer outcomes
-
Very strong generalized cancer resistance:
- Reduced immortalization and oncogenic transformation of fibroblasts in vitro.
- Markedly lower tumor incidence in vivo across multiple protocols. (Nature)
Longevity / aging
-
This is the key positive result:
- s-Arf/p53 mice show delayed appearance of aging phenotypes (better neuromuscular performance, preserved dermal thickness, reduced lipofuscin/oxidative damage markers, etc.). (Nature)
- Median lifespan is significantly increased (roughly on the order of ~15–20% in the main cohorts), and this persists even if you censor out all mice that died of cancer. Maximum lifespan is not clearly extended. (Nature)
-
Later work by Carrasco-García et al. dissected this further, showing that increased Arf/p53 dosage preserves stem-cell function, delays stem-cell exhaustion, and extends lifespan, again without obvious trade-off in degenerative pathology. (PMC)
Bottom line:
This is the one clear engineered mouse model where extra p53 copy number (together with extra Arf) → strong tumor suppression plus modestly extended lifespan and delayed aging, provided regulation is left intact.
2. Hyperactive or truncated p53: cancer-resistant but short-lived
These models don’t add extra gene copies, but they are crucial for interpreting the “more p53” question.
2.1 Truncated / hypermorphic p53 (p53+/m; p44 etc.)
-
Tyner et al. and Maier et al. created mice expressing truncated, hyperactive p53 isoforms (e.g., p44) or hypermorphic alleles (p53+/m). (ScienceDirect)
-
Phenotype:
- Very strong tumor resistance.
- But also premature aging: kyphosis, osteoporosis, reduced body weight, organ atrophy, stem-cell exhaustion, and shortened lifespan. (ScienceDirect)
These models are often cited as evidence that p53 activation is pro-aging, but the later “super p53” and “s-Arf/p53” work shows that this is a regulation problem, not an in-principle trade-off: when p53 is chronically and inappropriately active, you age faster; when it is only more responsive to damage but otherwise normally controlled, you can get cancer resistance without an obvious lifespan cost, and sometimes with a modest benefit.
3. Non-mouse mammals with extra TP53 copies
Here we mostly have comparative / natural experiments, not engineered animals.
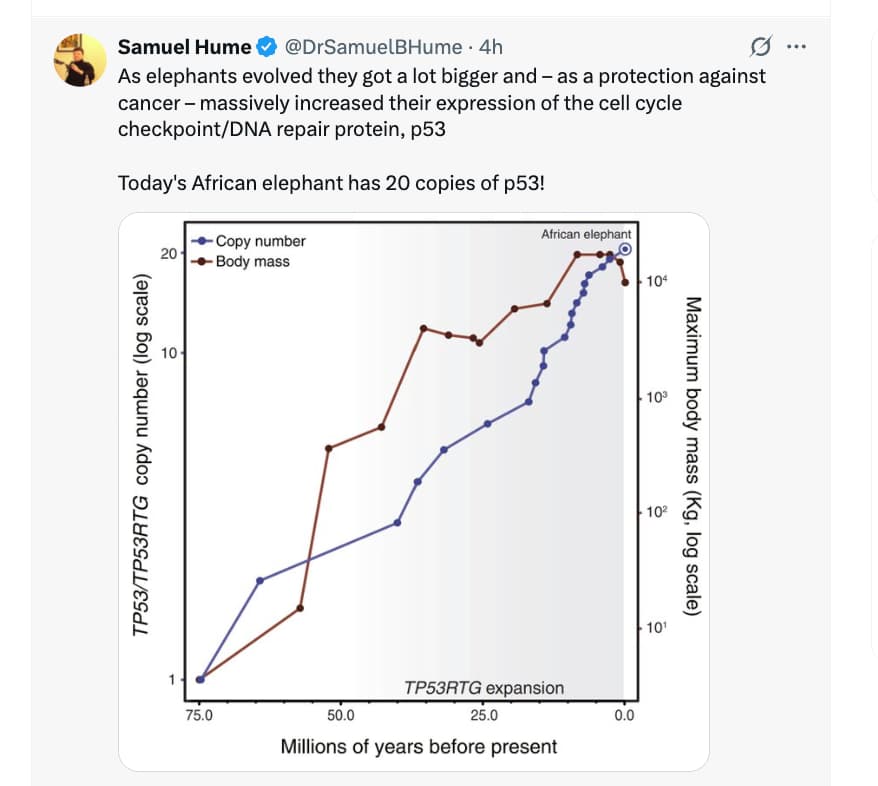
3.1 Elephants: 20 copies of TP53
-
African elephants have ~20 copies of TP53 (mostly retrogenes), versus 1 canonical TP53 gene in humans. Several of these retrogenes are expressed and contribute to a hyper-sensitive p53 signaling pathway. (PMC)
-
Elephant cells show:
- Much stronger p53-dependent apoptosis after DNA damage.
- A much lower estimated lifetime cancer mortality (~5%) compared with humans (~11–25%), despite many more cells and long lifespans—one proposed resolution to Peto’s paradox. (EvolutionMedicine)
This is consistent with the mouse “super-p53” story: increased p53 gene copy number, tightly regulated, seems compatible with long lifespan and low cancer risk. But in elephants this is an evolutionary outcome, not an engineered intervention, and you can’t cleanly decouple p53 from many other co-evolved traits.
To date there are no published reports of deliberately adding extra TP53 copies to other large mammals (dogs, primates, etc.) with lifespan readouts; work is mainly in mice plus comparative genomics in elephants, whales, naked mole-rats, etc. (PMC)
4. Condensed comparison table
| Model | Genetic change | p53 regulation | Cancer effect | Longevity / aging effect |
|---|---|---|---|---|
| Super p53 mouse (García-Cao 2002) | Extra wild-type Trp53 locus (BAC), 3 copies total | Native promoter; p53 only rises under stress | Strong reduction in spontaneous tumors; enhanced DNA-damage response | Normal aging; no clear lifespan extension (PMC) |
| High-p53 via low Mdm2 (Mendrysa 2006) | Diminished Mdm2 expression → chronically higher p53 | Still under endogenous Trp53 promoter; feedback relaxed | Marked cancer resistance | Normal lifespan; no overt progeria (PubMed) |
| Super Ink4a/Arf (Matheu 2004) | Extra Ink4a/Arf locus | Native regulation | Increased cancer resistance | Normal aging and lifespan (arep.med.harvard.edu) |
| s-Arf/p53 (Matheu 2007) | Extra Trp53 + extra Ink4a/Arf | Native regulation for both | Very strong, broad cancer resistance | Delayed aging, reduced molecular damage, modestly ↑ median lifespan, max lifespan unchanged (Nature) |
| Hyperactive p53 (p53+/m, p44, etc.) | Mutant / truncated alleles → chronically hyperactive p53 | Largely deregulated, stress-independent | High cancer resistance | Premature aging and shortened lifespan (ScienceDirect) |
| Elephant (natural) | ~20 TP53 copies (retrogenes + canonical) | Co-evolved, highly regulated system | Very low lifetime cancer mortality vs body size | Long lifespan with low cancer; not an engineered test but strong circumstantial support (PMC) |
5. How to interpret this for “cancer risk vs longevity”
What is clearly shown
-
Yes, extra p53 copies in mammals can lower cancer risk.
- In mice, an extra wild-type Trp53 copy gives robust tumor resistance. (PMC)
- In elephants, many TP53 copies correlate with unusually low cancer mortality. (EvolutionMedicine)
-
Extra p53 alone does not reliably extend lifespan in mice.
- Super-p53 and high-p53 models age normally; they don’t clearly live longer. (Science)
-
Combined Arf + p53 extra dosage, under normal regulation, can modestly extend lifespan and delay aging.
- s-Arf/p53 mice: higher median lifespan, delayed functional and molecular aging markers, and effect persists when you remove cancer deaths from the analysis. (Nature)
-
If p53 is made too active or misregulated, the trade-off flips: cancer resistance + faster aging + shorter life.
- Hyperactive p53 mutant/isoform models show that chronic stress-independent p53 activity is deleterious to stem cells and tissue maintenance. (ScienceDirect)
I wanted to dig deeper into CIRBP, so:
My Prompt:
Summarize and provide links to, the research on CIRBP cold induced RNA binding protein as it relates to health and longevity of mammals
CGPT5.1 Response:
CIRBP is clearly important for stress resilience in mammals; its role in longevity per se is promising but still mostly indirect, with one direct in vivo lifespan experiment (in flies, not mammals) and strong circumstantial data from bowhead whales.
Below is a structured summary plus links.
1. What is CIRBP?
Cold-inducible RNA-binding protein (CIRBP / CIRP / hnRNP A18) is an stress-responsive RBP induced by mild hypothermia, hypoxia, UV, and other cellular stresses. It
- Binds 3′UTRs of target mRNAs and modulates mRNA stability and translation.
- Participates in liquid–liquid phase separation and stress granule dynamics.
- Regulates apoptosis, metabolism, DNA damage responses, circadian timing, and cell-death pathways (e.g., ferroptosis).
Comprehensive recent review (good entry point):
-
Rana 2024 – “Unraveling the intricacies of cold-inducible RNA-binding protein” – broad survey of CIRBP in stress responses, metabolism, neuroprotection, cancer, and inflammation.
 PMC full text
PMC full text
2. CIRBP, bowhead whales, and DNA repair–linked longevity
The strongest longevity-relevant data are from the bowhead whale:
- Bowhead whales (~200–250-year lifespan) show ~100-fold higher CIRBP expression in fibroblasts and multiple tissues compared with other mammals.
- Bowhead CIRBP overexpressed in human cells:
- Enhances non-homologous end joining and homologous recombination DNA repair.
- Reduces micronuclei formation and promotes DNA end protection.
- Bowhead CIRBP overexpressed in Drosophila:
- Extends lifespan and improves resistance to irradiation.
Key paper:
-
Firsanov et al. 2025 – “Evidence for improved DNA repair in long-lived bowhead whales” (Nature) Nature article
Interpretation:
- This is the most direct link between a CIRBP ortholog and lifespan extension (but in flies, not mammals).
- In bowheads, high CIRBP appears to be a longevity-enabling adaptation via genome maintenance, plausibly driven by cold Arctic habitat.
Knowledge gaps:
- No mammalian experiment yet showing that CIRBP manipulation alone extends mammalian lifespan .
- We do not know the safe window for CIRBP up-regulation in humans given its cancer-context-dependent roles.
3. CIRBP in mammalian metabolism, stress resistance, and aging
8. How strong is the link between CIRBP and mammalian longevity?
Direct evidence:
- None showing that altering CIRBP alone extends lifespan in mammals.
- One in vivo lifespan study in Drosophila using bowhead CIRBP shows lifespan extension and improved irradiation resistance.
Indirect / circumstantial evidence in mammals:
- Bowhead whales: very high CIRBP expression is tightly associated with:
- Enhanced DNA repair.
- Low mutation rates.
- Exceptional lifespan and cancer resistance.
- Rodent/cell models: CIRBP repeatedly:
- Protects neurons, cardiomyocytes, and other cells from hypoxia, oxidative stress, ferroptosis, and amyloid toxicity .
- Supports mitochondrial function and mitophagy .
- Modulates glucose metabolism, feeding, and energy homeostasis under cold/fasting.
- Integrates circadian and temperature signals, which are known to impact aging trajectories.
Counter-vailing evidence / risks:
- CIRBP can promote tumorigenesis and chemoresistance in some cancers.
- Expression patterns change with age and cold adaptation in tissue-specific ways; we do not know whether restoring “youthful” CIRBP levels is net beneficial or harmful.
Bottom line:
CIRBP is emerging as a central stress-response / genome-maintenance node that plausibly contributes to longevity in cold-adapted, long-lived species (bowhead whales). However:
- In mammals, causal evidence for lifespan extension is currently lacking .
- Any translational strategy that chronically boosts CIRBP would require careful cancer-risk stratification and tissue-specific control.